
Debunkers have occasionally brushed aside the sea serpent phenomenon as "just" being reports of oarfish, giant squid, frilled sharks, et cetera; I highly question how many (if any) reports these casual debunkers have actually read. The public idea of sea serpents is apparently of a mythological entity rather than something that actually has some interesting reports. Oudemans and Heuvelmans were early researchers who wound up collecting hundreds of reports, some of which cannot easily be written off as hoaxes or known phenomenon. I should note that I'm by no means a credulous believer in cryptozoology, just because something can't be explained doesn't mean that it is proof of something unknown. The possibility of existence is raised because of some reports but the only way to confirm existence is through physical remains. This is a very odd state of being, seemingly real but maybe not, and is possibly the reason why people clump into believers and non-believers. I'll attempt to take a skeptical (i.e. critical) approach to the subject, in my opinion the most fair way of dealing with it.
With only one possible exception, the only plausible evidence for sea serpents comes anecdotal reports. So what merit can anecdotal evidence have? After dealing with historical giant snake reports, I'm suggesting that any pre-science "evidence" be treated as a curiosity and nothing more. Even the more recent reports tend to be vague and/or suspect most of the time, but once in a while there is a remarkable sighting that is detailed and appears credible. Like the Valhalla and the Daedalus sightings, for instance. More vague reports can possibly be used to indicate possible range for sea serpents, but I'd suggest even that may be stretching credibility. The better sea serpent sightings can be plausibly compared to the occasional reports of beaked whales not fitting any known species by scientists. In the past unknown forms were seen (Mesoplodon sp. A, Tropical Bottlenose Whale) and then classified (Mesoplodon peruvianus, Indopacetus pacificus) when physical evidence showed up or was examined more closely. Some species, such as the apparently distinctive M. traversii, have not been observed to date, so a low number of sightings alone isn't a good indication against existence. The fact that sea serpent sightings have only very rarely been observed by scientists and the fact that they seem to be highly derived from any known group means that the anecdotal evidence should be scrutinized much more than the ziphiids though.
After collecting and sieving through reports of anomalous marine animals, the next logical step is to try and determine exactly what was sighted. Anthonid Cornelis Oudemans proposed a single species of, uh, "viverrine"/otter/pinniped/Basilosaurus relative behind the sightings. The proposed family tree looked odd even for that time period (Basilosaurus as a Carnivoran!?!) and this marks a trend of researchers stretching palaentological evidence to fit their proposals. A few decades later Heuvelmans decided that Megophias megophias and its vast repertoire of surface behavior couldn't adequately explain the range of reports and proposed nine sea serpent types. Mammalian sea serpents dominated with the long necked (giant pinniped), merhorse (pinniped? archaeocete?), super otter (very primitive archaeocete), many humped (Basilosaurus-like archaeocete), and many finned (very derived archaeocete). I discussed the last one at some length and was baffled at how Heuvelmans came to the conclusion that he did with the information that he had - for one thing most reports never mentioned the titular characteristic! Oh and there were also multiple giant eels and eel-shaped fish, a giant crocodile-shaped reptile, a giant turtle (dropped), and a tadpole shaped giant yellow bellied something (dropped), here's a charming little illustration of the whole bunch. I don't think that Heuvelmans started off with preconceptions but his text hints that he had a fairly concrete idea of the types early on. Coleman and Huyghe recently attempted to clean up the system by combining the long necked and merhorse (waterhorse), the super eel and super otter and many humped (classic sea serpent) and basically kept the rest (mystery saurian, cryptic chelonian, great sea centipede). The team's bigfoot classification was problematic, but this classification seems more conservative and plausible (except for the giant beaver, egads). Not too much was added by the team and it appears that all of the problems with Heuvelmans got passed on, like finless many finned sea serpents. It really is just about time for someone to start this all again from the ground up.
The first indication of another classification system was "A Preliminary Evaluation of a Study of the Morphology, Behavior, Autoecology, and Habitat of Large, Unidentified Marine Animals, based on Recorded Field Observations" by Bruce A. Champagne in an online collection of Cryptozoology articles called Dracontology. Coleman and Huyghe cited the article but didn't discuss it, perhaps due to its preliminary nature. Aside from some excerpts getting posted on Cryptozoology.com, nothing else was published or mentioned for around six years. Late last year in the book Elementum Bestia, Mr. Champagne published "A Classification System for Large, Unidentified Marine Animals based on the Examinations of Reported Observations". It does seem to be very interesting indeed. There are now 1247 reports on file with much more data getting taken into account, such as water temperature, salinity, current et cetera. Although not mentioned in the report, I think recording data that the witnesses couldn't have been subjective about is an interesting way to make the reports seem more plausible (assuming there were patterns). Unfortunately, documentation of the reports is not done in the article - some of them were apparently odd enough that I'd like a way to track them down for myself. Perhaps it was beyond the scope of the article. Not all the reports were used anyways, and there was an established point system (+1 for a qualified observer, +1 for physical evidence, etc) that whittled the number down to 209. Quantifying quality of reports is subjective of course, but at least it is attempting to be more scientific than past incarnations. In fact, both articles are written in a style reminiscent of technical peer-reviewed literature.
The author is very mysterious and searches for past articles on marine biology and cryptozoology online have proved fruitless. If anybody has any more information on him (or if he's reading this!) please, do share.
Time to discuss the sea serpents themselves, and yes, there are more than 9.
Type 1: "Long-necked"
These are reports that, of course, are of long necked animals. Confusingly, other types have this characteristic (3, 4B), but presumably other characters took precedence. This type is somewhat comparable to the long-necked/merhorse/super-otter classification of Heuvelmans and the "waterhorse" category of Coleman and Huyghe. Unlike previous authors, it has been divided into two sub-types.
Type 1A:

Type 1B:

With only one possible exception, the only plausible evidence for sea serpents comes anecdotal reports. So what merit can anecdotal evidence have? After dealing with historical giant snake reports, I'm suggesting that any pre-science "evidence" be treated as a curiosity and nothing more. Even the more recent reports tend to be vague and/or suspect most of the time, but once in a while there is a remarkable sighting that is detailed and appears credible. Like the Valhalla and the Daedalus sightings, for instance. More vague reports can possibly be used to indicate possible range for sea serpents, but I'd suggest even that may be stretching credibility. The better sea serpent sightings can be plausibly compared to the occasional reports of beaked whales not fitting any known species by scientists. In the past unknown forms were seen (Mesoplodon sp. A, Tropical Bottlenose Whale) and then classified (Mesoplodon peruvianus, Indopacetus pacificus) when physical evidence showed up or was examined more closely. Some species, such as the apparently distinctive M. traversii, have not been observed to date, so a low number of sightings alone isn't a good indication against existence. The fact that sea serpent sightings have only very rarely been observed by scientists and the fact that they seem to be highly derived from any known group means that the anecdotal evidence should be scrutinized much more than the ziphiids though.
After collecting and sieving through reports of anomalous marine animals, the next logical step is to try and determine exactly what was sighted. Anthonid Cornelis Oudemans proposed a single species of, uh, "viverrine"/otter/pinniped/Basilosaurus relative behind the sightings. The proposed family tree looked odd even for that time period (Basilosaurus as a Carnivoran!?!) and this marks a trend of researchers stretching palaentological evidence to fit their proposals. A few decades later Heuvelmans decided that Megophias megophias and its vast repertoire of surface behavior couldn't adequately explain the range of reports and proposed nine sea serpent types. Mammalian sea serpents dominated with the long necked (giant pinniped), merhorse (pinniped? archaeocete?), super otter (very primitive archaeocete), many humped (Basilosaurus-like archaeocete), and many finned (very derived archaeocete). I discussed the last one at some length and was baffled at how Heuvelmans came to the conclusion that he did with the information that he had - for one thing most reports never mentioned the titular characteristic! Oh and there were also multiple giant eels and eel-shaped fish, a giant crocodile-shaped reptile, a giant turtle (dropped), and a tadpole shaped giant yellow bellied something (dropped), here's a charming little illustration of the whole bunch. I don't think that Heuvelmans started off with preconceptions but his text hints that he had a fairly concrete idea of the types early on. Coleman and Huyghe recently attempted to clean up the system by combining the long necked and merhorse (waterhorse), the super eel and super otter and many humped (classic sea serpent) and basically kept the rest (mystery saurian, cryptic chelonian, great sea centipede). The team's bigfoot classification was problematic, but this classification seems more conservative and plausible (except for the giant beaver, egads). Not too much was added by the team and it appears that all of the problems with Heuvelmans got passed on, like finless many finned sea serpents. It really is just about time for someone to start this all again from the ground up.
The first indication of another classification system was "A Preliminary Evaluation of a Study of the Morphology, Behavior, Autoecology, and Habitat of Large, Unidentified Marine Animals, based on Recorded Field Observations" by Bruce A. Champagne in an online collection of Cryptozoology articles called Dracontology. Coleman and Huyghe cited the article but didn't discuss it, perhaps due to its preliminary nature. Aside from some excerpts getting posted on Cryptozoology.com, nothing else was published or mentioned for around six years. Late last year in the book Elementum Bestia, Mr. Champagne published "A Classification System for Large, Unidentified Marine Animals based on the Examinations of Reported Observations". It does seem to be very interesting indeed. There are now 1247 reports on file with much more data getting taken into account, such as water temperature, salinity, current et cetera. Although not mentioned in the report, I think recording data that the witnesses couldn't have been subjective about is an interesting way to make the reports seem more plausible (assuming there were patterns). Unfortunately, documentation of the reports is not done in the article - some of them were apparently odd enough that I'd like a way to track them down for myself. Perhaps it was beyond the scope of the article. Not all the reports were used anyways, and there was an established point system (+1 for a qualified observer, +1 for physical evidence, etc) that whittled the number down to 209. Quantifying quality of reports is subjective of course, but at least it is attempting to be more scientific than past incarnations. In fact, both articles are written in a style reminiscent of technical peer-reviewed literature.
The author is very mysterious and searches for past articles on marine biology and cryptozoology online have proved fruitless. If anybody has any more information on him (or if he's reading this!) please, do share.
Time to discuss the sea serpents themselves, and yes, there are more than 9.
Type 1: "Long-necked"
These are reports that, of course, are of long necked animals. Confusingly, other types have this characteristic (3, 4B), but presumably other characters took precedence. This type is somewhat comparable to the long-necked/merhorse/super-otter classification of Heuvelmans and the "waterhorse" category of Coleman and Huyghe. Unlike previous authors, it has been divided into two sub-types.
Type 1A:

This "long necked" is primarily distinguished by a head of the same or slightly smaller diameter than the neck. Type 1As are reported worldwide, but appear most in boreal climate zones. They aren't even limited to salt water and have apparently been sighted several kilometers inland in fresh water, possibly to breed. Champagne also suggests that this type is a pinniped and a relatively large one at 2.5-12 (9 avg) meters in reported length. Given peoples' tendency to exaggerate, I'd suggest that this type could fall within the mass range of pinnipeds. The proposition of a long necked and tailed pinniped raises a lot of questions. Pinniped necks actually aren't longer than a dog's ('cept Acrophoca see Darren of course) and tend to be immensely thick to boot. Pinnipeds have very short tails, and the Coleman/Huyghe book suggested that reports of a long tail are due to the rear limbs. The superficial plesiosaur or elasmosaur-like body coupled with a pinniped-style flexible neck makes this type quite unique and would presumably indicate an unknown niche. The idea of a pinniped being fully adapted to a marine life and taking on a new form doesn't seem too outlandish, and at least this type resembles common sightings. The lack of resemblance to anything in the fossil record is still a major problem of course.
Type 1B:

This "type" is only known from 5 sightings in the North Atlantic and is distinguished by a head larger in diameter than the neck. It is supposedly much larger (17 meters+) than the 1A and displays more "primitive" characteristics and different behaviors (frequently associates with cetaceans, etc). Oh, these illustrations are ones that I did a while back, so you'll see I chose to portray it as a more robust "1A" type animal as opposed to another lineage of long-necked creature. Limbs were never observed and only inferred to exist by presumed relations. The proposed anatomy of this type is even stranger than the 1A, and I don't know what to think of a massive head on a long neck. Judging by the lack of sightings or apparently much detail, I'm suggesting that future analyses will probably just absorb these sightings into the "1A" or maybe "type 3" classification. Ah, to lump or to split, the eternal question.
Type 2: Eel-like
Animals described as just that, eel-like. Only about half of the reports were detailed enough to classify, and the three sub-types specified seem very divergent. The first two types have no analogy in Heuvelmans, and the last one (exemplified by the Daedalus sighting) is a bit super-eel or yellow belly-ish.
Type 2A: Although not outright mentioned, the reported details of this type match up the Tresco encounter, which Heuvelmans rejected due to its outlandish nature. Apparently there have been similar, more subdued sighting of a similar nature in the western Atlantic. Coleman and Huyghe noted that Mangiacopra once had a classification with an apparently similar sea serpent, but further information is lacking. This type is distinguished by a squarish head and scales. The head supposedly has pendulous lips, mucous emanating from the mouth, and either horns or tusks. The compliment of fins is confusing, it has variously noted as having a dorsal fin (once) as well as fan-like pectoral and/or anal (?!) fins. I portrayed it as a limbed sea snake-like animal with a very thick body (8-18% the total length). The reports average 6-9 meters with one 30 meter report (the Tresco). This type seems very...confused...and the idea of a reptilian walrus analogy off the Eastern coast of the US and Canada is quite a bit out there. I'd like to know the location of the other reports and how many there even were and if alternate explanations are possible (hoaxes?).
Although not outright mentioned, the reported details of this type match up the Tresco encounter, which Heuvelmans rejected due to its outlandish nature. Apparently there have been similar, more subdued sighting of a similar nature in the western Atlantic. Coleman and Huyghe noted that Mangiacopra once had a classification with an apparently similar sea serpent, but further information is lacking. This type is distinguished by a squarish head and scales. The head supposedly has pendulous lips, mucous emanating from the mouth, and either horns or tusks. The compliment of fins is confusing, it has variously noted as having a dorsal fin (once) as well as fan-like pectoral and/or anal (?!) fins. I portrayed it as a limbed sea snake-like animal with a very thick body (8-18% the total length). The reports average 6-9 meters with one 30 meter report (the Tresco). This type seems very...confused...and the idea of a reptilian walrus analogy off the Eastern coast of the US and Canada is quite a bit out there. I'd like to know the location of the other reports and how many there even were and if alternate explanations are possible (hoaxes?).
Although not outright mentioned, the reported details of this type match up the Tresco encounter, which Heuvelmans rejected due to its outlandish nature. Apparently there have been similar, more subdued sighting of a similar nature in the western Atlantic. Coleman and Huyghe noted that Mangiacopra once had a classification with an apparently similar sea serpent, but further information is lacking. This type is distinguished by a squarish head and scales. The head supposedly has pendulous lips, mucous emanating from the mouth, and either horns or tusks. The compliment of fins is confusing, it has variously noted as having a dorsal fin (once) as well as fan-like pectoral and/or anal (?!) fins. I portrayed it as a limbed sea snake-like animal with a very thick body (8-18% the total length). The reports average 6-9 meters with one 30 meter report (the Tresco). This type seems very...confused...and the idea of a reptilian walrus analogy off the Eastern coast of the US and Canada is quite a bit out there. I'd like to know the location of the other reports and how many there even were and if alternate explanations are possible (hoaxes?).Type 2B:

A much, much more mundane and believable "sea serpent", this actually appears to be an unknown beaked whale of some sort distinguished (from other type 2s) by a pointed head. One report mentioned an ambiguous structure behind the head (a "mane"), but this animal is suggested (by the author, not in the article) to have a series of bumps on the spine instead. Once again, I'd be curious to have the reports available because it would be interesting to know if the witnesses simply observed a known species of large (7.5-9 m) beaked whale and didn't note the dorsal fin. The type 3 sea serpent supposedly has a mane and series of humps as well. Sightings were rather uncommon (once every 40 years) and it doesn't seem like this type has much merit. It was seen in both the Atlantic and Pacific and hasn't been reported in over 80 years. What a shame, an unknown beaked whale (even one with a series of humps replacing a dorsal fin) is a fairly plausible cryptid by anyone's standards.
Type 2C:
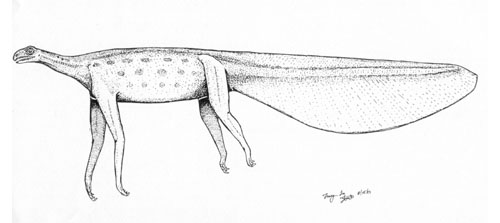
 The last type of "eel-like" animal, this time distinguished by a blunt "cow-like" head. The head is reported as being quite large proportionally (14-33% of the total length), which seems a bit jarring with the Daedalus illustration. It is also described with a tapering tail (making it tadpole shaped?). This is also a proportionally rather thick (10-16% total length) and long (18-24 m) animal with a rigid body and no reported appendages. Like the Daedalus sighting, it just isn't too clear how this animal is propelling itself. This type has been observed in the Atlantic and Indian Oceans as well as the South China Sea. It hasn't been observed in 70 years either. The mouth was described as sub-terminal, hard to see when closed, but overall large and with sharp teeth so Champagne hypothesized that this was a zooplankton feeder. As for classification, he proposes that this is a reptilian animal, and I'm not too clear on why. Apparently this is an aggressive tail slapper like a lepidosaur, although where it derived from I don't have a clue. I portrayed it as a very massive sort of sea snake or sea snake-like reptile.
The last type of "eel-like" animal, this time distinguished by a blunt "cow-like" head. The head is reported as being quite large proportionally (14-33% of the total length), which seems a bit jarring with the Daedalus illustration. It is also described with a tapering tail (making it tadpole shaped?). This is also a proportionally rather thick (10-16% total length) and long (18-24 m) animal with a rigid body and no reported appendages. Like the Daedalus sighting, it just isn't too clear how this animal is propelling itself. This type has been observed in the Atlantic and Indian Oceans as well as the South China Sea. It hasn't been observed in 70 years either. The mouth was described as sub-terminal, hard to see when closed, but overall large and with sharp teeth so Champagne hypothesized that this was a zooplankton feeder. As for classification, he proposes that this is a reptilian animal, and I'm not too clear on why. Apparently this is an aggressive tail slapper like a lepidosaur, although where it derived from I don't have a clue. I portrayed it as a very massive sort of sea snake or sea snake-like reptile.Type 3: Multiple-humped (available: Here and Here)

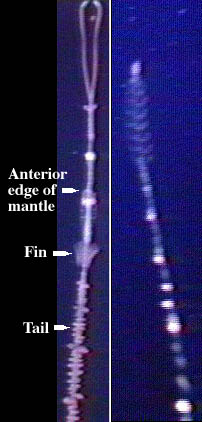
This is pretty much the sea serpent, it is responsible for about 1/3 of the sightings, is reported worldwide, and is synonymous with "Cadborosaurus" and others. Bruce wrote another article on the Clark brothers' video which he interpreted to be of this "type". It has been reported in areas from the open ocean to several kilometers inland in fresh water, and has been reported from estuaries with great frequency. Oh, and apparently it can venture over land. The anatomy of this type, despite it being so common, is absolutely baffling. The head is camel-like with a sub-terminal mouth, a spear-like tongue, prehensile lips, possible vomerine teeth, and maybe some sort of crest or a mane. The neck appears to be quite long (the foreflippers are located about 1/3 back) and the bilobate tail is plated and apparently capable of making sound. Oh yes, and apparently the body has yellow "spines" and it has been reported up to 60 meters in length (move over Amphicoelias!) with 10-20 meters average. If this reported animal actually does have all those variable characteristics, then I can't help but wonder if it is responsible for or confused for some of these other "types". The author doesn't specify the classification index, but appears to plump for a reptilian identity. I'd have no idea where this animal fits in classification-wise, presumably the only reptiles that could have adapted to vertical undulations would be archosaurs of some sort - even though crocodiles don't. The lack of evidence of anything remotely resembling this in the fossil record is a huge problem, but not outright disproof. Even if there are many good sightings and a weird carcass ("carcass"?), this type is still hugely unsettling.
Type 4: Sail finned
Another category of largely dissimilar animals sharing a vaguely similar feature. The first type has no analogy in the other systems, and the second (more common) type is somewhat comparable with the super eel and Valhalla sighting.
Type 4A: (Available: Here)
 Yet another "sea serpent" that appears to be a sort of beaked whale. I feel that this drawing came out the best, by the way. This animal is also from the northwest Atlantic Ocean and was unfortunately last seen in 1912. How many sea serpent types live off the Eastern US coast anyways? Apparently "Paxton 1995" discusses this animal, although it wasn't listed in the works cited. Despite the "crocodile-like" head and "scales" (warts? barnacles?), this type does seem to resemble a cetacean. Once again, I'm curious about the actual sightings to confirm (for myself) that this actually does resemble a large (9-21 m?) ziphiid with an unusually small head and large dorsal fin. A recently extinct beaked whale isn't too unusual by anyone's standards, and I'm sure that marine biologists would find the reports interesting.
Yet another "sea serpent" that appears to be a sort of beaked whale. I feel that this drawing came out the best, by the way. This animal is also from the northwest Atlantic Ocean and was unfortunately last seen in 1912. How many sea serpent types live off the Eastern US coast anyways? Apparently "Paxton 1995" discusses this animal, although it wasn't listed in the works cited. Despite the "crocodile-like" head and "scales" (warts? barnacles?), this type does seem to resemble a cetacean. Once again, I'm curious about the actual sightings to confirm (for myself) that this actually does resemble a large (9-21 m?) ziphiid with an unusually small head and large dorsal fin. A recently extinct beaked whale isn't too unusual by anyone's standards, and I'm sure that marine biologists would find the reports interesting.Type 4B: (Available: Here)
 Another very strange proposed animal, the author considers this "a transitional animal with reptilian and mammalian characteristics". The closest thing to an aquatic non-mammalian synapsid appears to be the Ophiacodontids - however they're very very old and don't show any obvious aquatic adaptations. The neck is apparently long and there are no appendages, so the animal supposedly moves by ostraciform swimming with the large dorsal fin. At a reported 4-26 meters long, this would be by far the largest animal using that locomotion and I wonder how effective it would be at such sizes. The head is of similar diameter to the neck and has a large mouth, lips, and "tendrils". The overall body shape, disregarding the inferred neck, does strongly suggest a fish of some kind, somewhat reminiscent of Derichthys. I think it is likely that this type will be included within another "type" in future classification.
Another very strange proposed animal, the author considers this "a transitional animal with reptilian and mammalian characteristics". The closest thing to an aquatic non-mammalian synapsid appears to be the Ophiacodontids - however they're very very old and don't show any obvious aquatic adaptations. The neck is apparently long and there are no appendages, so the animal supposedly moves by ostraciform swimming with the large dorsal fin. At a reported 4-26 meters long, this would be by far the largest animal using that locomotion and I wonder how effective it would be at such sizes. The head is of similar diameter to the neck and has a large mouth, lips, and "tendrils". The overall body shape, disregarding the inferred neck, does strongly suggest a fish of some kind, somewhat reminiscent of Derichthys. I think it is likely that this type will be included within another "type" in future classification.Type 5: Carapaced Animal
 Yet another animal described as being a "reptile-like mammal, or transitional/intermediate animal of unknown class" regardless of the resemblance to a turtle. The measurements (thickness 75-90% total length, thickness 60-75 total length) give it a fineness ration of a little over 1 and proved impossible to portray plausible, so something must have been wrong (flippers included?). Aside from the plated, shell-like carapace and long (!) wedge-shaped tail, the animal is occasionally described with oily hair, quills (!), and a seal-like head, probably resulting in the bizarre classification. I think if reports were selected differently a much more plausible looking animal could result, although plausible looking doesn't necessarily mean more likely.
Yet another animal described as being a "reptile-like mammal, or transitional/intermediate animal of unknown class" regardless of the resemblance to a turtle. The measurements (thickness 75-90% total length, thickness 60-75 total length) give it a fineness ration of a little over 1 and proved impossible to portray plausible, so something must have been wrong (flippers included?). Aside from the plated, shell-like carapace and long (!) wedge-shaped tail, the animal is occasionally described with oily hair, quills (!), and a seal-like head, probably resulting in the bizarre classification. I think if reports were selected differently a much more plausible looking animal could result, although plausible looking doesn't necessarily mean more likely.Type 6: Saurian (Available: Here)
 An animal with a crocodilian (occasionally seal-like) head ranging from 1-20 m in length from the (sigh) North Atlantic and Mediterranean. The occasional spots are the only new piece of information to this Marine Saurian/Mystery Saurian animal. Reports seem rare enough that this type will probably just remain some sort of curiosity.
An animal with a crocodilian (occasionally seal-like) head ranging from 1-20 m in length from the (sigh) North Atlantic and Mediterranean. The occasional spots are the only new piece of information to this Marine Saurian/Mystery Saurian animal. Reports seem rare enough that this type will probably just remain some sort of curiosity.Type 7: (Available: Here)
 I've talked about this animal far too much as it is. Apparently only 6 sightings were used, 3 of which are from carcasses (!) and one of which was photographed (!!!). That last bit of information should be major news, but there aren't any references to where it can be found. Even compared to the other improbable animals, this is a particularly troublesome one.
I've talked about this animal far too much as it is. Apparently only 6 sightings were used, 3 of which are from carcasses (!) and one of which was photographed (!!!). That last bit of information should be major news, but there aren't any references to where it can be found. Even compared to the other improbable animals, this is a particularly troublesome one.Type 8: Digited Animal (Available: Here)
 This type of "sea serpent" appears to be based off of the Canvey Island Monster to some degree, which was probably some relative of the anglerfish (poorly reported). The idea of an aquatic chameleon-like reptile has been proposed before in the case of Hypuronector, although it is now thought to be arboreal like the rest of the bizarro Avicephalans. This "type" can probably discarded due to suspected misidentifications. This really goes to show the importance of at least referencing the sightings used.
This type of "sea serpent" appears to be based off of the Canvey Island Monster to some degree, which was probably some relative of the anglerfish (poorly reported). The idea of an aquatic chameleon-like reptile has been proposed before in the case of Hypuronector, although it is now thought to be arboreal like the rest of the bizarro Avicephalans. This "type" can probably discarded due to suspected misidentifications. This really goes to show the importance of at least referencing the sightings used.Type 9: Snake-like (available: here)
A 6-18 meter long snake-like animal with a squared head and visible scales reported from the Atlantic. This seems very similar to the 2A sea serpent, although this wasn't mentioned. I made a preliminary drawing and sent it to Bruce, but he didn't give feedback and it didn't make it into his report.
Giant cephalopods were also mentioned but never discussed.
This does indeed seem to be a very interesting classification system. There are a lot of questions left to be answered, and I can't help but think that the scope of the article is somewhat responsible for that. I think a lot more discussion by the author would have been necessary rather than leaving it to the reader. There certainly does seem to be a lot of interesting information covered, and I think a better medium than a short (~30 page) article in a obscure book with no ISBN would be necessarily for the best airing. It still seems like a lot of people had not heard about this classification, hence me writing about it. Undoubtedly the story does not end here and somebody else in time will build on this and other system to make yet another one.
I don't know if we'll ever find a "sea serpent" in the ocean. Yes, yes, we're all aware how large and mysterious the oceans are, but an apparent unknown branch of aquatic, say, synapsids, stretches credibility past breaking point. I'd personally suggest that the most parsimonious explanation is that there is a single aberrant species which, combined with cultural preferences hoaxes and mundane phenomenon creates the sea serpent phenomenon. Well, that is assuming that there is an unknown species to begin with.
Well, I'm tired of Cryptozoology for now
-Cameron




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}